.png)

200813
Tolerância ou assimilação?
Nelson Vaz
Introdução
Dizer que a imunidade anti-infecciosa depende da produção de elementos defensivos, como anticorpos, é tão esclarecedor quanto dizer que o sal é salgado. No entanto, desde sua origem no século dezenove, a imunologia busca esclarecer este mecanismo de defesa, hoje entendido como se fosse um “sistema imune” altamente complexo. Nunca se considerou que esta proteção, que pode ser reforçada por vacinas, é um comentário que fazemos sobre o resultado de vários mecanismos do viver, e não um mecanismo defensivo destacado dos demais. É fácil confundir resultados da operação de mecanismos com os próprios mecanismos. A seleção natural, por exemplo, é entendida como um mecanismo importante no processo evolutivo, e que foi assim que Darwin a propôs. Mas a seleção natural pode ser vista como um resultado do viver, daquilo que Humberto Maturana e Jorge Mpodozis (2000) chamam de “deriva filogênica natural”. Seleção não é escolha, a natureza não escolhe; Darwin não a propôs desta maneira.
Derivas
O termo “deriva” não deve ser confundido com o acaso. Um barco à deriva no oceano não vai para qualquer lugar, não se desloca ao acaso: vai para um único lugar, determinado por um conjunto de acontecimentos que dependem de seu peso e tamanho, do vento, de ondas e de correntes marítimas — apenas não tem uma rota. Sua deriva continua enquanto ele flutua. A deriva termina quando o barco bate em um rochedo e afunda, ou as ondas o depositam em uma praia. Mas não podemos dizer que ele se aproximou desse rochedo ou dessa praia, porque ele não tinha uma rota em seu deslocamento. Seu trajeto era determinado instante a instante. Um exemplo similar é a epigênese, um processo importante no viver citado por Conrad Waddington (2012) um pioneiro em entender o que se passa com os seres vivos em seu viver.
Donald Forsdike disse que: “Para entender as ideias da imunológica é preciso conhecer sua história, mas para conhecer esta história, é preciso conhecer a imunologia”. Uma tautologia, como a do biscoito que vende muito porque está sempre fresquinho, e está sempre fresquinho porque vende muito.
Assim como o próprio viver, estes são processos dinâmicos, dependem do entendimento de mudanças. Mas como diz Mpodozis (2011) o que se conserva importa mais do que aquilo que varia:
“Há plasticidade nos modos de desenvolver. Os caminhos do desenvolvimento têm plasticidade em todos os momentos, e isso é o que permite essa maravilhosa diversidade de linhagens de seres vivos. Mas o problema não é o que é plástico, e sim o que se conserva. Se a mudança é uma condição constitutiva do viver, então, como se conserva aquilo que se conserva?” (Jorge Mpodozis. Epígrafe em “Onde está o Organismo?” Vaz et al., 2011), (Florianópolis, Editora UFSC)
Mudança e conservação
Esta preocupação com aquilo que se conserva não chegou à imunologia, ainda. A produção de elementos vistos como defensivos, como anticorpos, ou linfócitos ativados são mudanças em células consideradas inertes antes de serem “estimuladas” pelo antígeno. Aquilo que se conhece como “memória imunológica” e a memória em si mesma é algo que se conserva. As vacinas funcionariam porque, supostamente, criam uma “memória” do contato com um dado agente infeccioso contido na vacina em forma enfraquecida (atenuada). Quando o corpo encontra novamente este agente, ele reage mais intensamente. A imunologia estuda esta memória. Mas é uma conservação episódica, específica para um dado micróbio ou vírus. E a proteção contra múltiplos micróbios e vírus, portanto, dependeria de múltiplas vacinas, aplicadas de preferência na infância.
O ser vivo conserva a si mesmo enquanto muda continuamente sua estrutura molecular e celular. Usualmente, se considera isto tão complexo que não se pode abordar nem descrever. Mas sabemos que os seres vivos são entidades autônomas, estão entregues a si próprios, se auto-constroem e se auto-mantêm, um processo que Maturana chama de autopoiese (auto-criação/manutenção) (Maturana, 2002).
A autopoiese é o estado padrão (default) dos seres vivos. Em todos os momento de seu viver, sem exceção, os seres vivos, conservam a si mesmos como sistemas autopoiéticos moleculares; mudam continuamente de estrutura molecular, trocam moléculas (e também células) por outras que lhes são equivalentes, mas mantêm invariante sua organização autopoiética. Estrutura e organização são termos úteis para discutir a mudança e a conservação expressa pelos seres vivos.
Tolerância imunológica
O termo tolerância se refere a uma negação protelada. É um termo inadequado, mas se tornou importante desde cedo na história da imunologia. Na virada do século vinte, Ehrlich mostrou que o corpo pode formar anticorpos não apenas contra micróbios e toxinas, mas contra praticamente tudo o que lhe é estranho, variando de proteínas de plantas a hemácias de outro indivíduo da mesma espécie. Isto expandiu enormemente os horizontes da imunologia para além dos micróbios e da medicina. Ehrlich apontou também uma grande lacuna nesta reatividade: o corpo não forma, aparentemente, anticorpos contra seus próprios componentes; exibe o que ele chamou de horror autotoxicus, uma aversão a se auto-agredir — algo que Ehrlich considerava totalmente sem sentido, profundamente disteleológico. Para ele, fazia sentido admitir que o próprio corpo se ignora, imunologicamente.
Talvez por sua complexidade, o problema foi deixado de lado por meio século. Em 1949, ainda em seu trabalho como virologista, MacFarlane Burnet reviveu este problema com o nome de tolerância imunológica (Burnet and Fenner, 1949). Ele apontava o trabalho de Owen (1945) sobre gêmeos bovinos, que nascem como quimeras genéticas, pois contêm células um do outro durante toda a sua vida. A placenta gemelar em bovinos permite a mistura de sangue entre os fetos e os fetos recebem células tronco um do outro. Organismos vertebrados adultos rejeitam transplantes de órgãos e tecidos e outros indivíduos da mesma espécie (transplantes alogênicos), mas os dados de Owen mostram que o organismo pode viver em paz (tolerar) com células de outro organismo — desde que esta mistura ocorra na vida fetal. Burnet tentou estabelecer experimentalmente a tolerância imunológica em embriões de galinha. Seus experimentos falharam, mas alguns anos depois Medawar e seus companheiros teriam mais sorte.
Tolerância alogênica específica
A produção da tolerância imunológica no laboratório foi o primeiro passo a indicar que a atividade imunológica um problema muito mais complexo que a produção de anticorpos anti-microbianos. A invenção empírica de vacinas e a formação de anticorpos são os fatos que formaram a base da imunologia contemporânea, e surgiram antes de que a complexidade da atividade linfocitária fosse sequer esboçada. Mas toda esta complexidade foi sendo adicionada à ideia da formação de anticorpos específicos sem modifica-la em sua essência.
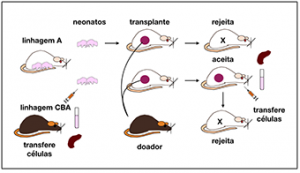
Nos anos 1950, Medawar e colaboradores mostraram que é possível gerar artificialmente a tolerância específica a transplantes de pele em camundongos geneticamente homogêneos (isogênicos). Eles expuseram embriões (ou recém-natos) de camundongos de uma linhagem a células de outra linhagem e assim os tornaram tolerantes a transplantes da linhagem doadora. Medawar sugeriu que esta tolerância dependia da ausência de linfócitos reativos com o transplante (linfócitos alo-reativos). O pequeno texto que eles publicaram (Billingham, Brent and Medawar, 1953), mudaria para sempre a face da imunologia. Medawar e Burnet receberam o prêmio Nobel de 1960 e, em seu discurso no banquete, Burnet declarou estes experimentos eram como fundadores de uma “nova imunologia” (Burnet, 1961).
Fig.1 Medawar et al. injetaram células da linhagem CBA em neonatos da Linhagem A, que, quando adultos, recebiam sem rejeitar transplantes de CBA. Transferidos para A tolerantes portando transplante, linfócitos de A colhidos de animal que rejeitou transplante CBA
induzem a rejeição.
Os dados de Billingham, Brent e Medawar (1953) sugerem que a tolerância alogênica se deva à inibição (ou destruição) de linfócitos especificamente reativos a alo-antígenos. Isto seria formaria a base da teoria de seleção clonal proposta por Burnet (1957) alguns anos depois.
Tolerância oral
Os antígenos mais comuns são proteínas formadas por outros seres vivos. A atenção estava incialmente voltada para componentes de micróbios, vírus e parasitas, mas a grande massa de material antigênico à qual o organismo é continuamente exposto vem dos alimentos e de produtos da microbiota nativa, que não é patogênica. O corpo forma anticorpos específicos para estes antígenos da dieta e da microbiota, mas não desenvolve uma “memória” destes contatos. Isto significa que os eventos imunológicos mais comuns e frequentes no viver saudável de todos os organismos não resultam em respostas imunes. Eles violam a regra admitida como universal para a reatividade imunológica! A assimilação de antígenos da dieta e da microbiota é uma espécie de “imunologia da paz” (assistir video em ) que nos mostra uma face oculta da memória imunológica (Vaz, 1981).
A tolerância a antígenos ingeridos como alimentos — chamada tolerância oral — foi registrada cedo na história da imunologia (Besredka, 1909; Wells, 1911; Chase, 1946), mas foi oculta na gaveta dos fenômenos curiosos e, curiosamente, nunca ocupou uma posição central na teoria imunológica. Nos anos 1970, este fenômeno foi “redescoberto” em vários laboratórios (Brandztaeg, 1996), inclusive no nosso (Vaz et al., 1977). Naquela época, mostramos que a tolerância oral pode ser transferida “adotivamente” para animais não-tolerantes pela transferência de linfócitos T (Richman et al., 1978). Isto mostra que se trata de um processo ativo e contradiz a ideia de que a tolerância depende da ausência de linfócitos reativos: não se pode transferir uma ausência.
Na mesma época em que literalmente “tropecei” na tolerância total (Vaz et al., 1977) aprendi a pensar sobre fenômenos “sistêmicos” com Francisco Varela (Vaz, 2011a) e entendi que a estabilidade (a conservação) dos sistemas naturais depende da manutenção de uma organização invariante (relações invariantes entre componentes), algo que Varela chamava de um “fechamento” (Varela and Johnson, 1976). Em um texto de 1981, distingo a visão tradicional da imunologia, que chamo de auto-desconhecimento, de uma visão sistêmica, na qual o “reconhecimento” de materiais externos se faz por con-fusão com componentes do próprio corpo (Vaz, 1981).
Assimilação e estabilidade
A investigação da tolerância oral passou a ser minha principal área de investigação tanto experimental quanto teórica. Vinte anos depois, constatamos que a reatividade residual que permanece em animais submetidos à tolerância oral, é robustamente estável, isto é, se conserva inalterada mesmo que o animal receba vários “boosters” com o antígeno (Verdolin et al., 2001). Também neste caso se trata de um fenômeno conservador e não da simples anulação de uma reatividade progressiva.
A reatividade imunológica do corpo a seus componentes também pode ser vista como um fenômeno conservador, e isto modifica todo um modo de ver. De um “estranhamento” a atividade imunológica passa a ser um problema de assimilação e de conservação de padrões de reatividade. A imunologia inteira está baseada na discriminação self-nonself, na separação entre o que pertence e o que não pertence ao organismo (Burnet, 1957). Mas, nas últimas décadas, tornou-se evidente que animais sadios formam anticorpos e ativam linfócitos T reativos a seus próprios componentes (Coutinho, Kazatchkine and Avrameas, 1995). Não se trata de uma “auto-reatividade fisiológica”: este termo pertence à visão do estranhamento. O que ocorre é a conservação de uma atividade auto-mantenedora/auto-construtora do organismo; a atividade imunológica é parte da conservação da organização autopoiética do organismo.
Ideias escandalosamente desafiadoras
Quem leu Maturana e não se escandalizou, não entendeu. Em seu “conhecer o conhecer” e ao redefinir o “viver na linguagem” dos seres humanos, ele define o conhecer como ação efetiva e podemos deixar de ver o pensar como algo abstrato. Tomo ideias de Maturana como base para uma redefinição dos problemas da imunologia. Uma aluna definiu o curso “Para entender a imunologia” como “escandalosamente desafiador”; fiquei feliz com o elogio e creio que o desafio está em se tornar “bilingue” em imunologia.
Não se trata de negar a imensa massa de conhecimento gerada pela imunologia experimental, mas sim de aceitar que as ideias na imunologia não viveram um progresso paralelo para explicar esta massa colossal de informações. Elas permanecem apoiadas em noções que hoje sabemos estarem equivocadas. A ideia de vacinar com “germes atenuados” e de “proteção” por anticorpos, geradas ainda no século dezenove, pertencem a uma imunologia pasteuriana que descreve um “estranhamento” em termos celulares e moleculares cada vez mais detalhados, mas são impossíveis de interpretar.
Sem exceção, as teorias imunológicas são teorias sobre linfócitos e anticorpos e não sobre um aspecto da atividade do organismo inteiro. Não sabemos inventar novas vacinas e provavelmente não há um mecanismo básico de proteção que possa ser aplicado a todas as doenças infecciosas. Não há cura para as doenças alérgicas e autoimunes que se tornam cada vez mais frequentes. Não há métodos seguros de transplantação de tecidos e órgãos ou de evitar complicações da gravidez.
Em contraste com exemplos de que a atividade imunológica pode rejeitar um câncer, o entendimento de como isto ocorre é ainda muito precário e a ideia premiada com o Nobel de que o segredo está em “inibir a regulação” de respostas imunes ao tumor, não é uma solução. Assim como no tratamento de doenças autoimunes por anticorpos monoclonais, ou, seu oposto, isto é, com doses maciças de imunoglobulinas (IVIg), o tratamento imunológico do câncer tem sucesso esporádicos e ainda oferece altos riscos de ser ele mesmo patogênico.
Com exceção da teoria da rede idiotípica (Jerne, 1974) todas a teorias imunológicas buscam explicar respostas imunes a estímulos antigênicos e sua regulação. Mas o organismo não requer estímulos agir e sua operação espontânea, fisiológica, tem sido ignorada pela imunologia. A relutância em abandonar o esquema estímulo-resposta-regulação deriva em parte do desconhecimento de uma biologia de sistemas. O que se passa com o observador humano em seu observar é o maior dos problemas e o imunologista não tem sido visto como um observador. Tenho insistido em que as observações imunológicas são específicas, mas que os os linfócitos e anticorpos, objetos destas observações, não são (Vaz, 2011b).
Bibliografia:
Besredka, A. (1909). De l'anaphylaxie. Sixiéme memoire de l'anapjylaxie lactique. Ann.Inst.Pasteur, 23, 166-174.
Brandtzaeg, P. (1996). History of oral tolerance and mucosal immunology.
Ann. N. Y. Acad. Sci., 778 ((Feb 13)), 1-26.
Burnet, M. F., & Fenner, C. (1949). The Production of Antibodies.
Melbourne: McMillan.
Billingham, R. E., Brent, L., & Medawar, P. B. (1953). Actively acquired tolerance of foreign cells. Nature, 172, 603-606. doi:http://dx.doi.org/10.1038/172603a0
Burnet, M. F. (1959). The Clonal Selection Theory of Immunity. Nashville,TE: The Vanderbilt and Cambridge Univ.Press.
Burnet, F. M. (1961). Immunological Recognition of Self (Nobel Lecture - 1960). Science, 133, 307-311.
Chase, M. (1946). Inhibition of experimental drug allergy by prior feeding of sensitizing agents. Proc.Soc.Exp.Biol.&Med., 61, 257-262.
Coutinho, A., Kazatchkine, M. D., & Avrameas, S. (1995). Natural autoantibodies. Curr Opin Immunol, 7(6), 812-818.
Jerne, N. K. (1974). Towards a network theory of the immune system. Ann. Immunol., 125C, 373-392.
Maturana, H., & Mpodozis, J. (2000). The origin of species by means of natural drift. Revista Chilena de Historia Natural 73:261-310 (2000), 73, 261-310.
Maturana, H. (2002). Autopoiesis, structural coupling and cognition: a history of these and other notions in the biology of cognition. Cybernetics & Human Knowing, 9(3-4), 5-34.
Owen, R. D. (1945). Immunogenetic consequences of vascular anastomoses between bovine twins. Science, 102, 400-401.
Richman, L. K., Chiller, J. M., Brown, W. R., Hanson, D. G., & Vaz, N. M. (1978). Enterically-induced immunological tolerance. I. Induction of suppressor T cells by ingragastric administration of soluble proteins. J.Immunol., 212, 2429-2435.
Vaz, N. M., Maia, L. C. S., Hanson, D. G., & Lynch, J. M. (1977). Inhibition of homocitotropic antibody response in adult mice by previous feeding of the specific antigen. J. Allergy Clin. Immunol., 60, 110. doi:http://dx.doi.org/10.1016/0091-6749(77)90035-5
Vaz, N. M. (1981). A face oculta da memória imunológica.
Ciência e Cultura, 33, 1444-1447.
Vaz, N. M. (1990). Construir a identidade. Contribuição de um imunologista.
Tese de doutorado, Deptº Bioquimica e Imunologia, ICB, UFMG, Belo Horizonte.
Vaz, N. M., Faria, A. M. C., Verdolin, B. A., & Carvalho, C. R. (1997). Immaturity, ageing and oral tolerance. Scand. J. Immunol., 46, 225-229.
Vaz, N. M. (2011a). Francisco Varela and the Immunological Self.
Systems Research and Behavioral Science, 28, 696–703. doi:DOI: 10.1002/sres.1126
Vaz, N. M. (2011b). The specificity of immunological observations.
Constructivist Foundations, 6(3), 334-351.
Verdolin, B. A., Ficker, S. M., Faria, A. M. C., Vaz, N. M., & Carvalho, C. R. (2001). Stabilization of serum antibody responses triggered by initial mucosal contact with the antigen independently of oral tolerance induction. Braz. J. Biol. Med. Res., 34(2), 211-219.
Waddington, C. H. (2012). The epigenotype. 1942.
Int J Epidemiol, 41(1), 10-13. doi:10.1093/ije/dyr184
Wells, H. G. (1911). Studies on the chemistry of anaphylaxis.III. Experiments with isolated proteins, specially those of the hen's egg. J.Inf.Dis., 9, 147-171.




- Apaixonados por Imunologia
- Comunicado
- Conteúdo Publicitário
- Curso
- Dept. Imunologia Clínica
- Dia da Imunologia
- Dia Internacional da Imunologia
- Divulgação científica
- Edital
- Especial
- Especial Dia da Imunologia
- Especial Doença de Chagas
- Evento
- Eventos
- Exposição COVID-19 da SBI
- História da Imunologia no Brasil
- Homenagem
- Immuno 2018
- Immuno2019
- Immuno2021
- Immuno2022
- Immuno2023
- Immuno2025
- Immuno2026
- IMMUNOLAC
- Immunometabolism2022
- Imune
- Imune - o podcast da SBI
- ImunoWebinar
- INCT Imuno
- Institucional
- IUIS
- Luto
- NeuroImmunology 2024
- Nota
- Nota Técnica
- Notícia
- o podcast da SBI
- Oportunidades
- Outros
- Parecer Científico
- Pesquisa
- Pint of Science 2019
- Pint of Science 2020
- Política Científica
- Sars-CoV-2
- SBI 50 ANOS
- SBI.ImunoTalks
- Sem categoria
- Simpósio
- SNCT 2020
- SNCT 2022
- Webinar
- WORKSHOP


