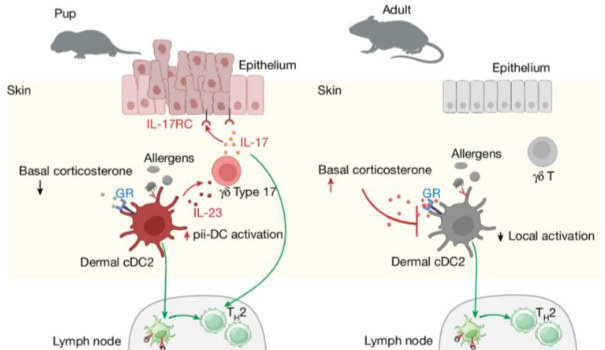
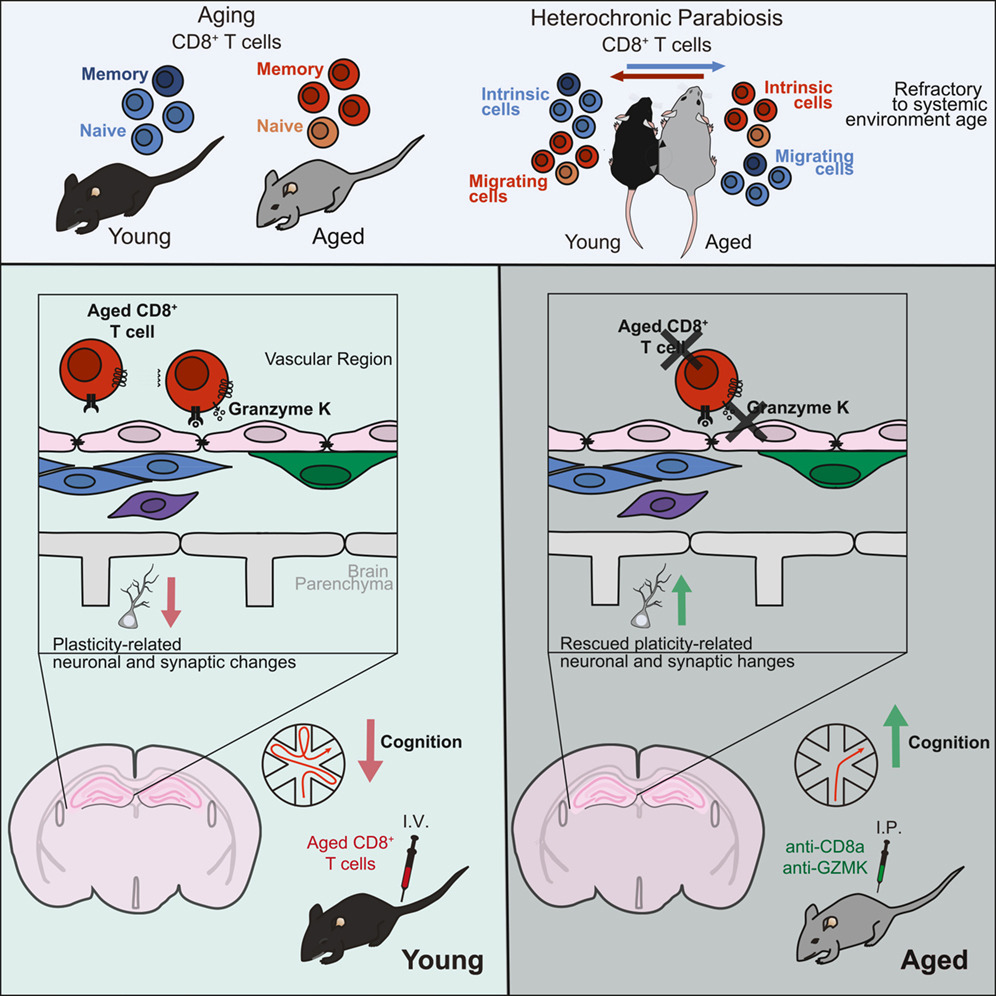
.png)

Muitos artigos na imunologia atual são tecnicamente tão sofisticados que tornam a leitura da seção “material e métodos” quase indecifrável, exceto para sub-especialistas. Quando vejo estudantes torturados por este problema, lembro-me de uma frase notável de Gregory Bateson, um dos gigantes intelectuais do século passado:
“…(M)uitas vezes na Ciência, e sempre na Arte, não se sabe
o que os problemas eram até depois que eles estejam resolvidos.”
A multidão de componentes moleculares e celulares já caracterizados na imunologia nos faz perguntar sobre o problema do qual eles, supostamente, são a solução encontrada pelos seres vivos em seu processo evolutivo.
Em sua teoria de seleção natural da produção de anticorpos, Jerne descreve a formação de uma enorme diversidade de imunoglobulinas, como um fenômeno natural, espontâneo, que não requer estimulação antigênica, e chamou estas globulinas de “anticorpos naturais” (Jerne, 1955). Esta foi uma constatação experimental e não uma conjectura sobre a natureza da atividade imunológica. Mais tarde se referiria a esta observação como “meu testamento”, isto é, sua declaração mais importante (Soderqvist, 2003/170).
Para que serviriam tantas moléculas diferentes? A qual pergunta esta diversidade quase inacreditável de anticorpos responde? A resposta usual diz que o organismo antecipa o encontro com agentes patogênicos, prevê o futuro. A imensa diversidade de anticorpos apontada por Jerne seria necessária para enfrentar uma diversidade comparável de materiais antigênicos que podem invadir o organismo, principalmente como componentes de vírus, micróbios e parasitas. Os anticorpos naturais nos protegeriam antecipadamente de invasões do corpo por antígenos.
A teoria de Jerne (1955) foi logo substituída pela teoria de Burnet (1957) que colocava a origem dos anticorpos naturais em clones de linfócitos, cada um dos quais forma um tipo de anticorpo. Mas a imensa diversidade de anticorpos que permitiria ao organismo antecipar qualquer invasor tornava inevitável a ocorrência de auto-anticorpos, isto é, de clones de linfócitos auto-reativos. Burnet, assim como Ehrlich meio século antes, achava que esta ideia desafiava o senso comum, era fortemente disteleológica (Ehrlich, 1900). Não faria sentido que o corpo se auto-destruísse com auto-anticorpos; a natureza não cometeria este erro. Os clones auto-reativos devem ser, portanto, proibidos. E a teoria de Burnet propõe a ideia de auto-tolerância e prevê a existência de doenças “auto-imunes” se ela for violada.
Até hoje não se considerou seriamente que isto pode ter outras explicações. Mas é bem possível que a “auto-reatividade” não seja destrutiva. O termo “auto” já indica a existência de uma entidade e a ideia de um “self” imunológico talvez seja dispensável (Vaz and Varela, 1978).
A imensa diversidade de anticorpos caracterizada experimentalmente por Jerne (1951), que surgia como um processo espontâneo (“natural”), independentemente de estímulos antigênicos, foi o achado mais importante de Jerne, e embora preterido pela teoria clonal, o fazia reclamar, anos mais tarde: “Ainda acho que minha teoria — de 1955 — era melhor!”.
Jerne levou 20 anos para propor uma segunda possibilidade aberta pela diversidade espontânea exibida no sistema imune. Desde que são tão diversos e reagem com virtualmente tudo, os anticorpos não apenas reagem com componentes do corpo que os produz: eles também reagem uns com os outros! Todos os anticorpos são auto-anticorpos naturais (Coutinho, Kazatchkine and Avrameas, 1995).
Esta é uma ideia revolucionária. Os auto-anticorpos naturais e os clones que os produzem estão entrelaçados, surgem como uma rede complexa multiconectada. Como todos os anticorpos são auto-anticorpos, nenhum é! — a ideia de auto-reatividade perde seu sentido classificatório e é substituída por uma rede complexa de linfócitos, imunoglobulinas e uma variedade de células acessórias, que Jerne chamiu de rede idiotípica. A região variável de cada anticorpo reconhece e é reconhecida pelas regiões variáveis de outros anticorpos e linfócitos. A atividade imunológica tem como base uma rede idiotípica (Jerne, 1974) que pode ser vista como “auto-suficiente”- (Jerne, 1974b).
Isto parece desvituar a ideia de imunidade anti-infecciosa, de reconhecimento de materiais estranhos, que é a ideia fundadora da imunologia. Uma imunologia onde os linfócitos reagem com tudo e não há “nada estranho” — como poderia isto nos defender doenças infecciosas? Esta é uma pergunta que merece ser reformulada agora que sabemos que os micróbios são milhares de vezes mais frequentes e diversificados do que supúnhamos. Mas a rede idiotípica pode, sim, sugerir mecanismos que resultam na defesa anti-infecciosa com base no próprio entrelaçamento entre os linfócitos.
Imagens internas
É possível imaginar tipos de anti-anticorpos para um dado anticorpo (Ac1): ele pode ser “reconhecido” por um outro anticorpo, que vou chamar de Ac2-alfa, ou pode “reconhecer” um detalhe (epitopo) semelhante ao antígeno em um outro anticorpo (idiotopo) que vou chamar de Ac2-beta (Ac2 b). Este Ac2 b é como se fosse uma “imagem interna” do detalhe antigênico (epitopo). Imagine um anticorpo Ac1 que reage (sem danificar) com um componente da membrana celular e que reage também com um Ac2 b que é uma “imagem interna” deste componente da membrana. Se este componente da membrana por acaso funcionar como “receptor” para um vírus, a presença de Ac2 b no líquido extracelular que banha esta célula será uma proteção contra o vírus.

Generalizando, a presença no plasma e no líquido extracelular de imunoglobulinas que funcionam como “imagens internas” de componentes das membranas celulares, cria uma barreira para a ligação de substâncias estranhas ao corpo nestas membranas. Esta é uma proteção determinada estruturalmente, que não sendo “antecipatória”, resulta na proteção do corpo. É também um exemplo de respostas que são construídas antes que surja a pergunta à qual respondem.
Bibliografia
Bteson, G. (1972) "Double bind, 1969"
In Steps to an Ecology of Mind, Ballantine Books, New York, p 271.
Burnet, F. M. (1957). A modification of Jerne's theory of antibody production using the concept of clonal selection. Austr.J.Sci., 20, 67-69.
Coutinho, A., Kazatchkine, M. D., & Avrameas, S. (1995). Natural autoantibodies.
Curr Opin Immunol, 7(6), 812-818.
Ehrlich, P. (1900). On immunity, with special reference to cell life.
Proc.Roy.Soc. (London), 66(432), 424-448.
Jerne, N. K. (1951). A study of avidity based on rabbit skin responses to diphtheria toxin-antitoxin mixtures. Acta Pathol. Microbiol. Scand., 87 (Suppl. 1), 1-183.
Jerne, N. K. (1955). The natural selection theory of antibody formation. Proc. Natl. Acad. Sci. U.S.A., 41, 849-857. doi: http://dx.doi.org/10.1073/pnas.41.11.849
Jerne, N. K. (1974). Towards a network theory of the immune system. Ann. Immunol., 125C, 373-392.
Jerne, N. K. (1974 ). Network notions. Annual Report of the Basel Institute for Immunology, 1974,, BII Report < www.bii.ch>, 6.
Soderqvist, T. (2003). Science as autobiography. The troubled life of Niels Jerne. New York: Yale University Press, p.170.
Vaz, N. M., & Varela, F. G. (1978). Self and nonsense: an organism-centered approach to immunology. Med. Hypothesis, 4, 231-257. doi:http://dx.doi.org/10.1016/0306-9877(78)90005-1



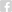

- Apaixonados por Imunologia
- Comunicado
- Conteúdo Publicitário
- Curso
- Dept. Imunologia Clínica
- Dia da Imunologia
- Dia Internacional da Imunologia
- Divulgação científica
- Edital
- Especial
- Especial Dia da Imunologia
- Especial Doença de Chagas
- Evento
- Eventos
- Exposição COVID-19 da SBI
- História da Imunologia no Brasil
- Homenagem
- Immuno 2018
- Immuno2019
- Immuno2021
- Immuno2022
- Immuno2023
- Immuno2025
- Immuno2026
- IMMUNOLAC
- Immunometabolism2022
- Imune
- Imune - o podcast da SBI
- ImunoWebinar
- INCT Imuno
- Institucional
- IUIS
- Luto
- NeuroImmunology 2024
- Nota
- Nota Técnica
- Notícia
- o podcast da SBI
- Oportunidades
- Outros
- Parecer Científico
- Pesquisa
- Pint of Science 2019
- Pint of Science 2020
- Política Científica
- Sars-CoV-2
- SBI 50 ANOS
- SBI.ImunoTalks
- Sem categoria
- Simpósio
- SNCT 2020
- SNCT 2022
- Webinar
- WORKSHOP


